Introdução aos princípios conexionistas
Por Rogério Perino de Oliveira Neves
Introdução
Próximo ao final do século XIX, Santiago Ramon y Cajal realizou
experimentos determinantes para a formação inicial do modelo conexionista,
concluindo que o cérebro era formado por células discretas, conectadas entre si
por ligações especiais, chamadas posteriormente de sinapses. Porém, a anatomia
do cérebro é estudada há séculos. Os antigos gregos já praticavam dissecação,
tendo realizado as primeiras observações e experimentos básicos no estudo do
sistema nervoso, dando início a vários dos ramos da ciência que estudam o
cérebro (neuroanatomia, neurofisiologia, psicofísica, psicologia, etc.).
Aqui
sintetizaremos brevemente alguns dos experimentos que foram significativos para
o ponto de vista conexionista, enfocando os mais recentes e algumas das
aplicações derivadas desse conhecimento.
Em
seguida, apresentamos uma breve introdução sobre o funcionamento dos neurônios,
células básicas que constituem o cérebro, suas várias formas e organizações em
redes.
Finalmente,
teremos uma síntese das tecnologias derivadas do estudo sistemático das redes
neurais, desde a criação de modelos de redes neurais artificiais para solução
de problemas específicos, até a tentativa de modelamento e simulação de um
sistema nervoso real, utilizando tecnologias digitais.
Toda
riqueza histórica do estudo da mente é retratada em detalhes em literaturas
específicas; aqui, retrataremos apenas eventos de maior notoriedade, deixando
outros que, embora não menos importantes, são menos necessários ao entendimento
introdutório da filosofia conexionista objetivado neste texto. O leitor que
desejar se aprofundar em algum dos assuntos cobertos poderá recorrer às
referências fornecidas na relação bibliográfica.
Por
muitos séculos persistiu a crença de que o cérebro abrigava um pequeno ser,
oculto na glândula pineal, onde Descartes acreditava ser “o sítio da alma”. Tal
ser era chamado de homúnculo e era responsável pelo controle e ações de todo o
corpo. O conhecimento adquirido ao longo dos séculos vem contribuindo para
descaracterização desse ser. Porém, muito ainda há para ser estudado e
explicado. A ciência nega a seus praticantes o direito de ignorar dados e
experimentos, mesmo que seus resultados, por vezes, entrem em conflito com
crenças advindas de bases sociais, culturais e religiosas. Devemos recorrer ao
passado, lembrando que muitas dessas crenças “dogmáticas” caíram por terra ao
longo dos séculos, com o avanço científico, o que ocasionou conflitos com
forças vigentes ao longo da história. Aqui trataremos apenas de fatos e dados
com embasamento científico, deixando discussões filosóficas e religiosas para
debates complementares.
N.Y.S.
Kiang ilustra o estudo do cérebro com uma observação singular:
“…
Neurofisiologia pode ser descrita como uma ciência rica em dados, mas pobre em
teoria”.
Experimentos significativos
Na
antiga Grécia há registro da prática da dissecação e das primeiras observações
registradas sobre o cérebro. Atribui-se a Hipócrates a observação: “… o
cérebro parece uma glândula, é branco e separado em pequenas massas, como as
glândulas são…”. Herófilo descreve os troncos nervosos como tubos que unem
as extremidades do corpo à medula espinhal e ao encéfalo. Posteriormente,
Galeno associa esta estrutura à transmissão da informação sensorial e motora.
Em
1644, Descartes forneceu uma descrição detalhada do arco reflexo, uma resposta
muscular a um estímulo doloroso. Em 1650, experimentos foram realizados pelo
holandês Swammerdam que, removendo um músculo da perna de uma rã, verificou que
este se contraia sempre que era perturbado ou irritado.
No
início do século XIX, Müller, considerado por muitos o “pai da fisiologia”,
escreveu um prognóstico negativista (típico dos cientistas daquela época)
dizendo: “…provavelmente nunca conseguiremos medir a velocidade da ação
nervosa, uma vez que não podemos comparar sua propagação em um espaço interno
como fazemos com a luz.”. Acreditava-se, até então, que os impulsos
nervosos propagavam-se com a velocidade da luz. Pouco durou tal pensamento. Em
1850, Hermman Ludwig Ferdinand von Helmholtz, pupilo de Müller e um dos mais
importantes cientistas do século XIX, mede a velocidade de propagação dos
estímulos nervosos, com um aparato inteiramente mecânico, obtendo o valor de
61m/s para os nervos sensoriais do homem, valor inteiramente consistente com os
obtidos com técnicas atuais. Mais tarde, Dubois-Reimond descobre a natureza
elétrica da ação nervosa e realiza medidas precisas da propagação dos estímulos
utilizando galvanômetros.
Ainda
no século XIX, o inglês Bell e o francês Megendie realizaram experimentos em
diversas partes do mecanismo sensorial, sintetizando-os em uma lei que, pela
primeira vez, estabelece uma taxonomia das funções realizadas no sistema
nervoso.
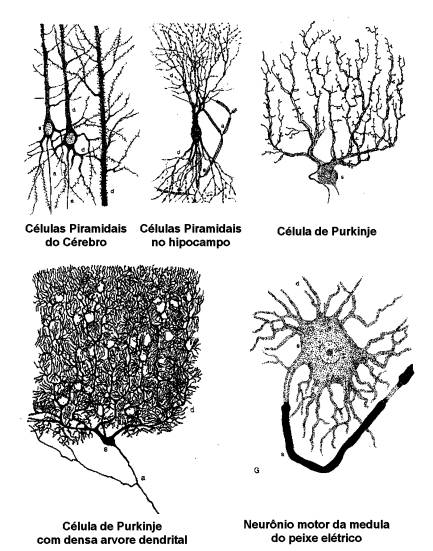
A
neuroanatomia celular, iniciada em 1837 por Purkyne, teve seu grande avanço com
trabalhos realizados pelo cientista espanhol Ramon y Cajal, que descrevem com
grande detalhe a taxonomia de vários neurônios do sistema nervoso e suas
respectivas árvores dendritais, assim como aglomerados dessas células, formando
as redes de processamento de informação no sistema nervoso.
Em
1906, o cientista inglês Sherrington publica “The Integrative Action of the
Nervous System”, que descreve experimentos feitos em animais, associando
diversas deficiências causadas por remoção de partes do córtex e medula
espinhal, criando os termos “neurônio” e “sinapse” para descrever a célula
nervosa e seus botões de contato. Em 1909, Brodman mapeia o córtex humano em 50
áreas por critérios cito-estruturais.
O
avanço mais significativo em eletroneurofisiologia se deu graças a Hudgkin e
Huxley, que modelaram em 1960 os processos biofísicos envolvidos na geração do
potencial de ação do neurônio, trabalho que lhes valeu o prêmio Nobel de
Fisiologia em 1963.
Desenvolveu-se,
ainda, a neurofisiologia funcional, onde certas funções primitivas do sistema
nervoso são associadas às áreas corticais específicas, mediante estudo
sistemático de pacientes lobotomizados, que tiveram de alguma maneira parte do
córtex avariado ou removido. Neste aspecto, ressalta-se o trabalho do
neurofisiologista Roger Sperry, que recebeu em 1981 o prêmio Nobel de
Fisiologia pelo seu trabalho.
Atualmente,
tecnologias como a microscopia eletrônica e técnicas modernas de mapeamento de
funções cerebrais, como tomografia computadorizada (CAT-Scan, PET-Scan) e
ressonância magnética (NMR), permitem a visualização do sistema nervoso em
pleno funcionamento, levando a um avanço rápido e eficiente das ciências que estudam
o sistema nervoso, deixando pouco espaço para o “homúnculo”.
O neurônio biológico
O
neurônio é constituído por um corpo somático, ou soma de onde derivam inúmeros
dendritos, formando uma árvore dendrital, e um único axônio, que pode
apresentar algumas ramificações. Sua taxonomia foi inicialmente estudada por
Ramón y Cajal, que descreve em detalhes vários tipos de neurônios encontrados
em sistemas nervosos. Sua extensão pode variar desde algumas centenas de
mícrons até alguns metros.
As
conexões entre os neurônios se dão por meios de sinapses, que podem ser
excitatórias ou inibitórias, dependendo do mecanismo e neurotransmissores
envolvidos nessa sinapse. O funcionamento do mecanismo sináptico é pouco
conhecido, porém, vem sendo estudado exaustivamente em vários ramos da ciência
(medicina, biofísica, biologia, etc.). Os dendritos se conectam aos axônios de
outras células, servindo como receptores de sinal, enquanto o axônio tem a
função de transmitir um pulso, dada a condição de disparo, conhecida como
“potencial de ação”. Na transmissão do impulso elétrico, a sinapse efetua a
troca de ions entre o citoplasma do axônio do neurônio transmissor e o dendrito
do neurônio receptor.
A
condição de disparo se dá quando o potencial da célula em relação ao meio
salino que a cerca atinge um determinado patamar. Esse patamar é conhecido como
limiar de disparo. Quando uma célula encontra-se com potencial positivo ela é
dita “polarizada”; é então acionado um mecanismo somático conhecido como
“bombas de íons” visando a diminuição da diferença de potencial na membrana até
o nível normal. Uma vez atingido o limiar de disparo, a célula é dita
hiperpolarizada, é disparado um único pulso no axônio, levando o potencial da
célula ao valor de despolarização absoluta, onde permanece por um período de
refração absoluta (Ta), onde a célula é incapaz de produzir outro potencial de
ação, não importando a intensidade da despolarização. Isto limita a freqüência
de operação do cérebro a 1/Ta. Posteriormente, há um período de refração
relativa (Tr), onde o potencial retorna assintoticamente ao valor normal.
Hudgkin
e Huxley foram os primeiros a estudar o potencial de ação, realizando
experimentos onde, além de obter valores precisos de tensão natural e de
disparo, ainda produziram reações em neurônios “in vitro”, aplicando uma tensão
nos dendritos, obtendo disparos induzidos de células removidas do sistema
nervoso.
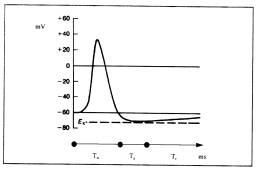
O
potencial de ação na membrana do axônio de um neurônio típico.
Redes neurais naturais e artificiais
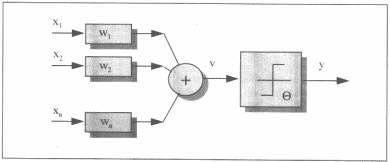
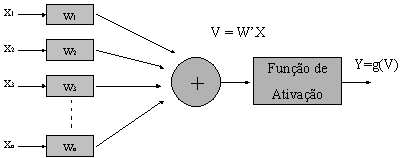
Uma
vez estabelecida a fisionomia e funcionamento do neurônio, podemos simplificar
sua representação considerando um sistema de n entradas com pesos distintos
aplicados, que, sendo x a soma ponderada,
dada uma função de ativação g(x), gera uma única saída como função da
entrada.
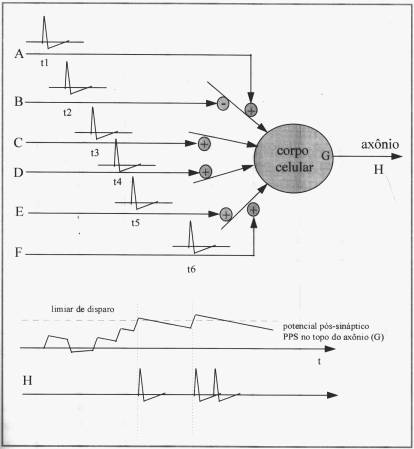
Modelo
simplificado do neurônio
Uma
vez determinada a função g(x), o comportamento desse neurônio é puramente
determinístico. A função g(x) do neurônio é conhecida e retratada na figura
abaixo.
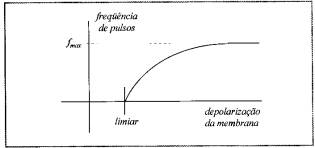
Resposta
neural: freqüência da despolarização dada a intensidade do estímulo
A
função característica do neurônio então se dá pela integração temporal-espacial
dos estímulos recebidos:
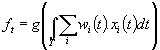
É aí
que o leitor atento pergunta: Então o cérebro nada mais é que uma máquina
composta de inúmeras unidades analógicas funcionais determinísticas? Por que,
então, não podemos determinar as ações de cada indivíduo? Ninguém poderia
responder a primeira pergunta sem confrontar filosofias e crenças
pré-estabelecidas. Já o comportamento não determinístico da mente, dada a
simplicidade de sua estrutura, remete à teoria de autômatas celulares, que
afirma que uma população de autômatas que implementam funções determinísticas
simples, quando combinados em estruturas complexas, podem produzir resultados
complexos e não determinísticos.
Para
efeitos de simplificação de modelos, a função resposta pode ser aproximada por
várias funções analíticas:
Funções
de ativação comumente empregadas em modelos de neurônios
Baseado
nestas aproximações e nos trabalhos realizados por Alan Turing e John von
Newman (que afirmavam que a álgebra booleana era a natureza essencial da
inteligência), Warren McCulloch, médico, filósofo, matemático e poeta,
juntamente com o estatístico Walter Pitts, publicaram um artigo no Bulletin of
Mathematical Biophysics com o título: “A Logical Calculus of the Ideas Immanent
in Nervous Activity”, que se tornou referência absoluta para a teoria das redes
neurais artificiais, sendo este tambem o primeiro paper escrito nessa área.
O
modelo de neurônio booleano era ingenuamente simples, exageradamente simples,
considerando toda a informação já disponível naquela época sobre o
comportamento elétrico da célula nervosa. Porém, baseado neste modelo,
McCulloch implementou uma série de funções, onde, eram atribuídas as entradas
um ganho arbitrário, gerando uma soma devidamente ponderada (ou subtração no
caso de sinapses inibitórias), resultando em uma única saída: pulso, no caso da
soma exceder um dado limiar, ou não-pulso, caso contrário.
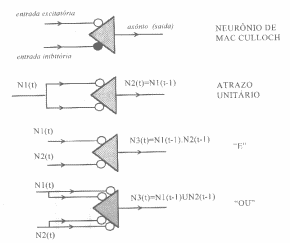
Algumas
funções booleanas implementadas por McCulloch
O
neurônio booleano nada mais é que um caso particular de um descriminador
linear, de entradas binárias que pode ser representado pela função:
![]()
A
função signal retorna –1 para parâmetros negativos e +1 para positivos,
e o teta representa o limiar de ativação.
Modelo
do neurônio como discriminador linear
Em
1950, Rosenblatt, na Universidade de Cornell, fez a primeira implementação
genuína das idéias de McCulloch, utilizando múltiplos neurônios do tipo
discriminadores lineares em rede de múltiplas camadas, experimento a qual deu o
nome de “perceptron”. O perceptron possui uma camada de entrada,
uma de saída, e uma ou mais camadas internas a entrada e saída, que são geralmente
referias como camadas ocultas.
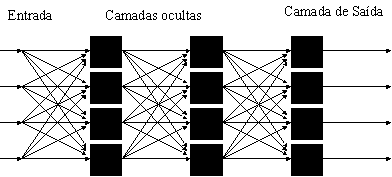
Modelo
do perceptron de Rosenblatt
A
primeira vista, o discriminador linear de Rosemblatt não passa de uma função
linear de n dimensões onde os pesos de cada conexão dados
arbitrariamente, e de fato, neste estado é de pouco uso. Porem, dado um
conjunto de exemplos com xi entradas e yi
saídas desejadas, este mecanismo se torna um poderoso reconhecedor de padrões,
que , como os similares biológicos, é capaz de aprender uma determinada função.
Utilizando-se de um processo de treinamento com o uso dos exemplos, é possível
alterar os pesos de cada conexão de forma a obter (na maioria dos casos) um
determinado conjunto de saída, dado um conjunto de entrada.
Baseado nesta teoria, Rosenblatt construiu um perceptron, o qual
batizou de mark1, onde um vetor de entrada de 400 dimensões advindo de uma
matriz de fotocélulas organizadas em matriz 20 x 20 pixels era colocado na
entrada, podendo reconhecer caracteres de um alfabeto, uma vez devidamente
treinado para este fim.

Padrões
de um alfabeto de dígitos 0-9, corretamente classificados por uma rede neural
O
treinamento visa a convergência da função para um limite que separe
corretamente grupos em um conjunto de n dimensões. Um modelo de treinamento
proposto por D. Hebb consiste nos seguintes passos:
![]()
![]()
O
parâmetro k é referido como taxa de aprendizado, geralmente é igualado a um,
porém, pode-se atingir uma convergência mais rapidamente modificando-o conforme
o conjunto de treinamento utilizado.
A
utilização do modelo acima é eficiente na maioria dos casos separáveis, e
mostra-se que a função converge para o conjunto desejado em poucas interações.
No caso de conjuntos não totalmente separáveis, ou que contenham
cross-sections, quanto maior o número de exemplos aplicados, menor será o erro
obtido nas classificações. Erros aceitáveis nestes casos situam-se entre 1% e
20% dependendo da natureza da intersecção dos conjuntos. Se os conjuntos forem
completamente superpostos, a rede jamais converge, e fica oscilando entre
valores intermediários do cruzamento.
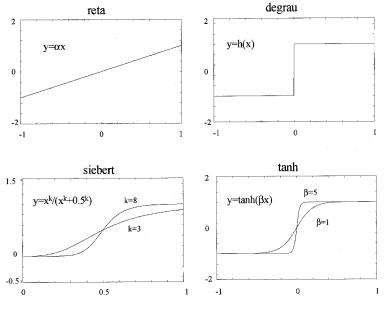
O
neurônio de Rummelhart, Hinton e Williams (RHW) foi o primeiro a empregar o
algorítimo de backpropagation, que resolvia algumas limitações fundamentais no
treinamento de redes complexas.
Neurônio
de Rummelhart, Hinton e Williams (RHW).
A retropropagação de erro (error
backpropagation) é um modelo de treinamento mais robusto e com álgebra mais
elaborada, atualmente o mais utilizado em redes neurais de múltiplas camadas,
uma vez que se demonstrou muito eficiente na maioria das aplicações
convencionais. A álgebra deste método é encontrada em Kovács (ver referência) e
outras literaturas específicas.
Redes
neurais atuais encontradas em aplicações comerciais e projetos aplicam as mais
diversas técnicas para minimizar os erros de classificação, aperfeiçoar o
conhecimento com erros detectados em tempo de execução (auto-adaptação), criar
discriminadores não lineares (redes neurais com base radial e multifuncionais)
e funções de ativação das mais esdrúxulas, de forma a obter um resultado
análogo, ou ao menos aproximado, ao deferido por um ser humano ordinário.
Alguns
exemplos de tecnologias comerciais que utilizam sistemas com redes neurais
artificiais são:
Reconhecedores
de escrita: Palm, handhelds, Pocket-PCs, etc.
Reconhecedores
de voz: Dragon, IBM-Via-Voice, Vocall.
Identificadores
de padrões visuais: Sistemas especialistas, satélites, radares, etc.
Sistemas de sensoriamento industrial: Para falhas mecânicas em linhas de
produção e montagem.
Sistemas
classificadores diversos.
Projetos
em inteligência artificial, vida artificial, ciências cognitivas, entre outros.
Finalizando
esta breve introdução aos princípios da teoria conexionista, seguem algumas
frases célebres conectadas de certa forma à ciência cognitiva num contexto mais
abrangente. Para refletir e expandir suas possibilidades neuro-sinápticas.
“Quando
eu disse ao caroço de laranja, que dentro dele dormia um laranjal inteirinho,
ele me olhou estupidamente incrédulo".
-
Hermógenes, filósofo e cientista grego.
“...
Mentes criativas são reconhecidas por resistir a todo tipo de mau treinamento”.
-
Anna Freud, psicanalista.
"Imaginação
é mais importante que inteligência".
-
Albert Einstein – Físico alemão.
"O
gênio fala muitas vezes mal e não sabe gramática. Mas transporta montanhas,
constrói cidades, estabelece estradas de ferro e telégrafos. Muitos homens
cultos falam e escrevem muito bem, mas são incapazes de construir e
criar".
-
Prentice Mulford.
"Aprender
com a experiência dos outros é menos penoso do que aprender com a
própria".
-
José Saramago - Escritor português.
"A
fábrica do futuro terá apenas dois operários: Um homem e um cachorro. A função
do homem será alimentar o cachorro, e a do cachorro será não deixar o homem tocar
nas máquinas".
- Walter Block - Autor de Defending the Undefendable
“...
Se você obedece a todas as regras, perde toda a diversão...”.
- Katherine Hepburn, atriz.
Bibliografia
Geoffrey M. Cooper – The Cell, ASM Press
Kovács,
Z. L. – O cérebro e sua mente, Edição
acadêmica
Kovács,
Z. L. – Redes neurais artificiais,
Edição acadêmica
Imagens
cedidas por Zsolt Kovács.
Rogério
Perinio de Oliveira Neves
http://www.lsi.usp.br/~rponeves